1. What are the basic principles of primer design?
The following principles of primer design are provided for your reference:
- It is recommended to design primers within the conserved regions of the template cDNA.
- Primer length should generally be between 15-30 bases.
- The GC content of the primer should be between 40%-60%, and the Tm value is recommended to be close to 72°C.
- The 3' end of the primer should avoid the third position of the codon.
- The 3' end of the primer should not be an A; T is recommended.
- Bases should be randomly distributed.
- Complementary sequences should not exist within a primer or between different primers.
- The ΔG values at the 5' end and the middle of the primer should be relatively high, while the ΔG value at the 3' end should be low.
- The 5' end of the primer can be modified, whereas the 3' end cannot be modified.
- The single strand of the amplification product should not form secondary structures.
- Primers must possess specificity.
2. What are the commonly used primer design softwares?
Commonly used softwares include Oligo 6 and Primer Premier 5.0. Primer design software is programmed based on general guidelines for primer design. In fact, the absolute key to the success of PCR amplification lies in reaction template preparation and reaction condition control. The disadvantage of primer design software is that it occasionally determines that no region of a specific gene meets the criteria for standard primers.
3. Can primer and probe sequences found in published literature be used directly?
Generally, sequences from international peer-reviewed journals are highly reliable and can be used directly. However, to ensure success, it is recommended to perform a BLAST search to verify the primer and probe sequences. Furthermore, analyzing the secondary structures and annealing temperatures of the primers and probes using primer design software will help you gain better control over the entire experiment.
4. How to calculate the Tm value of primers?
Definition of Tm value:
The DNA melting temperature (Tm) refers to the temperature at which half of the DNA double-helix structure is dissociated into single strands. In other words, it is the temperature during DNA denaturation when the ultraviolet absorbance reaches 50% of its maximum value.
MEBEP Tm value can be calculated using the following methods:
For primers that are 20mer or shorter, the Tm calculation formula is: Tm = 4°C(G+C) + 2°C(A+T). However, this formula is only applicable to primers with 14 to 20 bases. The Tm value is also affected by primer length, base composition, and the ionic strength of the buffer solution used.
For longer oligonucleotides, the Tm calculation formula is: Tm = 0.41(% of GC) – 675/L + 81.5
Note: L = total number of primer bases; % of GC = percentage of GC content = (Number of G and C bases / Total number of primer bases)
5. What are the common primer modifications?
| Modification | Description |
|---|---|
| Phosphorylation | 5' Phosphorylation can be used for linkers/adapters, cloning, gene construction, and ligase-catalyzed ligation reactions. 3' Phosphorylation is utilized in assays requiring resistance to 3' exonuclease digestion and is also used to block DNA polymerase-catalyzed DNA chain extension. |
| Biotin | Primer biotinylation can be applied in non-radioactive immunoassays for protein detection, intracellular chemical staining, cell separation, nucleic acid isolation, hybridization detection of specific DNA/RNA sequences, and ion channel conformational changes. |
| Digoxigenin | Digoxigenin is linked to the C5 position of uracil via an 11-atom spacer arm. Hybridized digoxigenin probes can be detected using anti-digoxigenin antibodies. Digoxigenin-labeled probes are widely used in various hybridization reactions, including Southern blotting (DNA-DNA), Northern blotting (DNA-RNA), Dot blotting, colony hybridization, in situ hybridization, and enzyme-linked immunosorbent assays (ELISA). |
| Internal Amino Modification | This is primarily achieved by adding a C6-dT aminolinker to a thymidine residue for internal modification. Post-modification, the amino group is positioned 10 atoms away from the main chain, which allows for further labeling and enzymatic ligation (e.g., alkaline phosphatase). Currently, dT-Dabcyl, dT-Biotin, and dT-Digoxigenin modifications mediated by internal amino modification are available. |
| 5' Amino Modification | Used to prepare functionalized oligonucleotides widely applied in DNA Microarrays and multiplexed diagnostic systems. Both 5' C6 amino modification and 5' C12 amino modification are available. The former is used to conjugate compounds that will not affect oligonucleotide function even when positioned closely, while the latter is used for attaching affinity purification groups and certain fluorescent labels, particularly when fluorescence might be quenched if positioned too close to the DNA strand. |
| 3' Amino Modification | Currently, 3' C6 amino modification is available. It is ideal for designing novel diagnostic probes and antisense nucleotides. For instance, the 5' end can be labeled with highly sensitive 32P or fluorescein while the 3' end is amino-modified for other conjugations. Additionally, 3' modification can inhibit 3' exonuclease digestion, making it useful for antisense experiments. |
| Thiol | 5'-Thiol modification is similar to amino modification in many aspects. Thiol groups can be used to attach various modifications, such as fluorescent labels and biotin. For example, thiol-linked fluorescent probes can be prepared in the presence of iodoacetic acid and maleimide derivatives. 5' Thiol modification mainly utilizes 5'-Thiol-Modifier C6-CE Phosphoramidite or Thiol-Modifier C6 S-S CE Phosphoramidite. After modification with the 5'-Thiol-Modifier C6-CE monomer, silver nitrate oxidation must be performed to remove the trityl protecting group, whereas the Thiol-Modifier C6 S-S CE monomer modification requires DTT to reduce the disulfide bond into a thiol group. |
| Spacer | Spacers provide necessary distance for oligonucleotide labeling to minimize interactions between the labeling group and the oligonucleotide. They are mainly used in DNA hairpin and double-stranded structure studies. The C3 spacer mimics the three-carbon interval between the 3' and 5' hydroxyl groups of ribose, or "replaces" an unknown base in a sequence. 3'-Spacer C3 introduces a 3' spacer arm to prevent 3' exonuclease and 3' polymerase activity. Spacer 18 is commonly used to introduce a strongly hydrophilic group. |
| Phosphorothioate (S-oligos) | Phosphorothioate-modified oligonucleotides are primarily used in antisense experiments to prevent degradation by nucleases. While full phosphorothioate modification is available, the Tm value of the oligonucleotide decreases as phosphorothioate bases increase. To mitigate this effect, phosphorothioate modification can be restricted to 2–5 bases at both ends of the primer, typically choosing 3 bases each at the 5' and 3' ends. |
| DeoxyUridine (dU) | Deoxyuracil can be inserted into oligonucleotides to raise the melting temperature of the duplex, thereby increasing its stability. Each replacement of deoxythymidine with deoxyuracil can increase the duplex melting temperature by 1.7°C. |
| DeoxyInosine (dI) | Deoxyinosine is a naturally occurring base. While not a true universal base, it forms relatively more stable mismatches when pairing with other bases compared to standard mismatches. The binding affinity of deoxyinosine with other bases follows the order of dI:dC > dI:dA > dI:dG > dI:dT. Under DNA polymerase catalysis, deoxyinosine preferentially pairs with dC. |
6. Why do modified primers have lower yields and higher prices than standard primers?
This is primarily because modified monomers have lower stability, require longer coupling times, and exhibit lower coupling efficiency, naturally leading to a lower final yield compared to standard primers. Furthermore, modified primers usually require PAGE or HPLC purification, which involves significant loss during the purification process. The raw materials used for modified primers can cost hundreds of times more than standard primer materials, which naturally results in a higher product price.
7. How should synthesized fluorescently labeled probes be stored?
Fluorescent probe storage methods are as follows:
- 1) Fluorescent probes must be stored away from light (protected from light).
- 2) Dry/lyophilized probes can be stored at -80°C for over a year. If unavailable, please store them at -20°C.
- 3) It is highly recommended to dissolve the probes in RNase-free TE buffer (pH 8.0) to achieve a more stable probe solution and a longer shelf life. Typically, reconstituted probes are prepared as a 100 pmol/μl stock solution and aliquoted into multiple tubes (each aliquot should withstand a maximum of 5 freeze-thaw cycles) for storage at -20°C. Before use, dilute the stock solution into a working solution (10 pmol/μl or 20 pmol/μl), and store the remaining portion at -20°C.
8. What are the common fluorescent dyes?
Fluorescent Dye Parameters
| Abbreviation | Full Name | Excitation Wavelength | Emission Wavelength | Color |
|---|---|---|---|---|
| 6-FAM | 6-carboxy-fluorescein | 494nm | 518nm | Green |
| TET | 5-tetrachloro-fluorescein | 521nm | 538nm | Orange |
| HEX | 5-hexachloro-fluorescein | 535nm | 553nm | Pink |
| TAMRA | tetramethyl-6-carboxyrhodamine | 560nm | 582nm | Rose |
| ROX | 6-carboxy-x-rhodamine | 587nm | 607nm | Red |
| Cy3 | Indodicarbocyanine | 552nm | 570nm | Red |
| Cy5 | Indodicarbocyanine | 643nm | 667nm | Violet |
9. What is the difference between FITC and FAM?
5-FAM, 6-FAM, and FITC are all fluorescein labels, where 5-FAM and 6-FAM are isomers of each other. Since their chromophores are all fluorescein, there is generally no difference in routine applications.
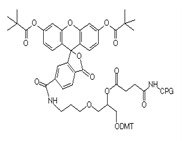
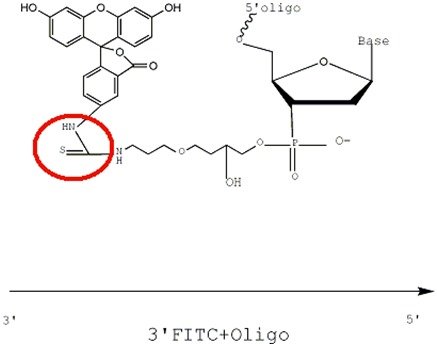
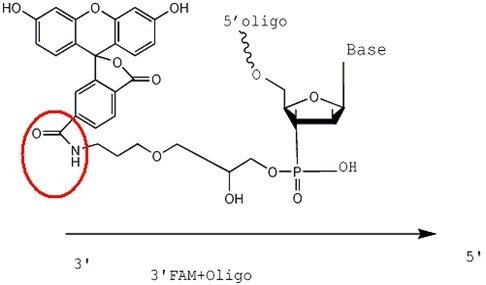
Structural formula of 3'FAM-CPG raw material
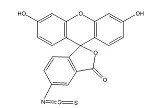
Structural formula of FITC raw material
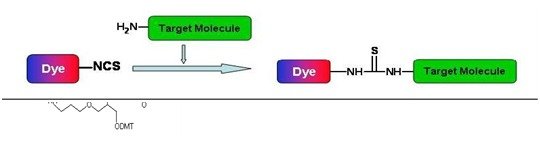
FAM is linked to the Oligo via an amide bond, whereas FITC is attached via a thiourea bond. The reaction process between -SCN on FITC and -NH2 proceeds as follows:
The figures below illustrate the effect after 3'FITC and 3'FAM are conjugated with the primer. The conjugation methods for 5'FAM and 5'FITC follow the same principle.
10. What are the differences in application among dual-labeled fluorescent probes with TAMRA, Eclipse, or BHQ series dyes as quenchers?
Dual-labeled fluorescent probes composed of quenchers like TAMRA, Eclipse, or BHQ series dyes are frequently utilized as hydrolysis probes (or TaqMan probes) in real-time quantitative PCR (qPCR) assays.
1) TAMRA is a fluorescent dye that emits fluorescence at higher wavelengths while quenching the reporter group. In contrast, Eclipse and the BHQ series are dark quenchers (non-fluorescent dyes) that emit no fluorescence of their own when quenching the reporter group. Consequently, they offer lower background fluorescence and higher detection sensitivity than TAMRA.
2) TAMRA features a narrow absorption spectrum, limiting the types of compatible reporter groups. Eclipse offers a broader absorption range (390 nm–625 nm) capable of quenching a wide array of reporter groups, including FAM, HEX, TAMRA, and ROX. The BHQ series dyes provide an even wider cumulative absorption spectrum spanning from 430 nm to the near-infrared, accommodating a broader selection of reporters including Cy3 and Cy5. Therefore, Eclipse or BHQ series dyes are ideal for designing dual-labeled fluorescent probe sets for multiplex PCR.
11. What are the basic principles of TaqMan probe design?
The following principles are provided for your reference:
- 1) The TaqMan probe should be positioned as close to the amplification primers as possible (amplicon size 50-150 bp), but must not overlap with the primers.
- 2) The probe length should generally be 18-40mer.
- 3) The G-C content should be maintained around 40-80%.
- 4) Avoid runs of identical bases, especially avoiding GGGG or longer poly-G stretches.
- 5) Avoid using a G base at the 5' end of the probe.
- 6) Incorporate a higher proportion of C bases when possible.
- 7) The annealing temperature (Tm) should be controlled around 68-70°C.
12. Do modification labels affect OD value measurements?
Certain fluorophores also absorb light at 260 nm, such as FAM, HEX, TAMRA, and TET. While other groups may also exhibit absorbance at 260 nm, they are excluded from calculations if their exact spectral data are unpublished.
13. What is phosphorylation and how does it work?
5' Phosphorylation is introduced to the 5' sugar ring of the primer via β-cyanoethyl chemistry rather than to the final base itself. 3' Phosphate is attached directly to the solid support medium, meaning the base couples to it during the first cycle of synthesis. The 3' phosphorylation modification serves to block extension by polymerases.
14. What is the difference between phosphorothioate (S-oligos) primers and standard primers?
S-oligos are formed when a non-bridging oxygen atom in the phosphodiester bond between single nucleotides of an oligonucleotide is replaced by a sulfur atom. This modification is introduced during the coupling step rather than post-synthesis. Once a base is added, it undergoes coupling modification. The phosphate group between two bases can be converted into a double-bonded "S" (using a sulfurizing reagent) instead of a standard double-bonded "O" (using an iodine solution); any base linked via a phosphodiester bond can undergo this modification.
However, the absolute 3'-terminal base cannot undergo phosphorothioate modification because when it is cleaved from the solid support, the phosphate ester bond is no longer present, leaving a 3'-terminal OH group. 5' modifications on S-oligos can be provided, allowing clients to customize fluorescein labeling on S-oligos. Both S-oligos and standard oligonucleotides migrate identically on PAGE gels.
15. What is the principle of amino modification, and what are the precautions?
5' Aminolinker (C6) is added to the 5' sugar ring of the primer in the form of a phosphoramidite via β-cyanoethyl chemistry during the final step of the synthesis cycle, rather than being attached to the ultimate base itself.
The standard coupling efficiency for 5' amino modification is >95%. The amino group displays no absorbance at 260 nm, though it absorbs at 210 nm. Its presence cannot be verified via gel electrophoresis (either agarose or polyacrylamide).
3' Aminolinker (C7) is only compatible with specific 5' modifications, such as FAM, HEX, TET, Fluorescein, Biotin, Amine, and phosphate. Other 5' modifications (such as Alexa dyes) require an amino group to attach to the oligonucleotide, making it impossible to prevent the dye from reacting with the 3' amino group as well.
Amino modification represents an uncomplicated and cost-effective approach. Any variant of amino modification is viable, with the 5'-terminal C6 type yielding excellent performance.
16. What are the differences among HEX, TET, and 6-FAM?
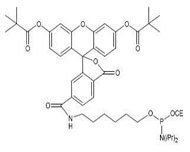
FAM, HEX, and TET are incorporated as phosphoramidites via β-cyanoethyl chemistry at the end of the synthesis cycle, attaching to the sugar ring at the 5' terminus of the primer rather than to the terminal base itself. They bind covalently to the final 5' sugar ring via a phosphodiester bond.
Solution color presentation: HEX appears pink, FAM appears yellow, and TET appears orange.
Oligonucleotides carrying these modifications cannot undergo subsequent phosphorothioate modification.
| MW | Emission (nm) | Excitation (Abs) Max (nm) | Extinction (at max) | Extinction (at 260) | |
|---|---|---|---|---|---|
| FAM | 554.5 | 517 | 494 | 74,850 | 20,960 |
| TET | 692.2 | 538 | 522 | 85,553 | 16,255 |
| HEX | 761.1 | 553 | 535 | 95,698 | 31,580 |
Note: Molar extinction coefficients are determined at the maximum excitation wavelength. Subtle variations in pH or buffer composition may influence these experimental data.
Primer Synthesis and Purification
Primer Synthesis, Storage, and Usage
Primer Design and Modification